GENÉTICA DEL PEZ LUCHADOR DE SIAM, BETTA SPLENDENS
HENRY M. WALLBRUNN
Departamento de Biología, Universidad de Florida, Gainesville, Florida
Primera edición 13 Marzo, 1957
El Betta Splendens, más comúnmente conocido como el Pez Luchador de Siam ha sido popular en los acuarios de Europa del Oeste y América alrededor de 35 años. Los antecedentes de su domesticación y consecuente cría selectiva se remontan a su introducción en el Oeste desde hace unos 60 o 70 años. Selección por agresividad, largas aletas y colores brillantes en este largo periodo han producido un número de fenotipos, ninguno de los cuales es muy similar a la forma salvaje de aletas cortas de los ríos lentos y arrozales inundados de Tailandia (Smith 1945).
El Betta de acuario se advierte por sus colores brillantes y variados. Éstos son producidos por tres pigmentos, luteina (amarillo), erythropterina (rojo) y melanina (negro) (Goddrich, Hill y Arrick 1941) y fragmentando la luz a través de pequeños cristales hexagonales (Goodrich y Mercer 1934) dando azul acero, azul o verde. Cada tipo de pigmento esta contenido en un tipo de célula distinto, xanthóforas conteniendo el amarillo, erythroforas rojo y melanóforas negro. No hay dos cromatóforas conteniendo dos pigmentos como es la xanthoerythóforas del Xiphophorus helleri. Las células reflectantes responsables del azul iridiscente y el verde iridiscente son conocidas como iridióforos o guanáforas y son más superficiales que las otras conocidas como cromatóforas.
Desde los gránulos de pigmentación podrían dispersarse bastante en varios pseudóforos bifurcados o agrupados en pequeños puntos en el centro de las cromatóforas, el color de un único pez podría variar entre un amplio rango de tonos y podría hacerlo en unos pocos segundos. Los peces enfermos o bajo estrés tienen una gama débil y uniforme sin dispersión de pigmentos; donde, sin embargo, cortejando o peleando con otro pez están en el extremo opuesto.
En los Bettas parece que hay también una continuidad completa en los tonos desde el marrón claro al negro puro. La misma condición existe respecto al rojo y también respecto a los colores producidos por los iridióforos. Su infinita variedad de fenotipos hace difícil la separación de las puestas en clases, pero el examen de grandes puestas debe mostrar el criterio y método correcto de examinar que la aparente continuidad cae en unos cuantos bloques largos dentro de los cuales, sin embargo, todavía parecen continuos. La diferencia entre los grupos largos puede trazarse hasta la segregación de unos pocos genes que llamaremos mayores.
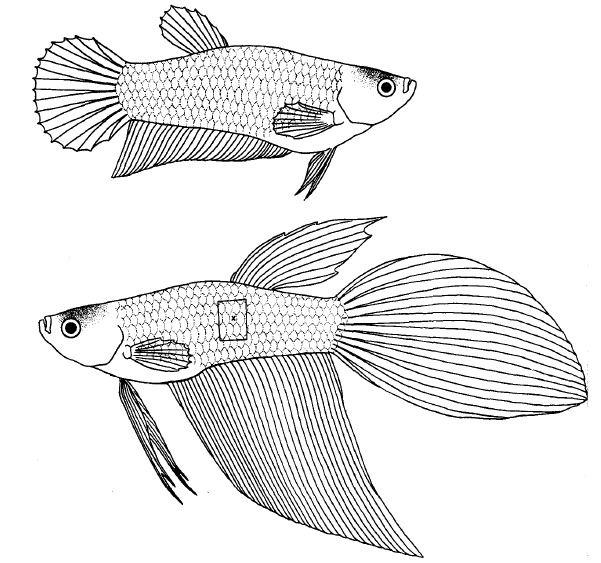
fig 1. Betta Splendens doméstico. Hembra arriba, macho abajo. En el rectángulo resaltado en la fig 2, x corresponde a las escamas contenidas en el área estándar para el recuento de melanóforos.
La aparente continuidad dentro de los grupos largos es proporcionada con toda probabilidad por factores menores pero lamentablemente ésto no ha sido testeado adecuadamente.
Material y métodos
Si la luz es reflejada por los iridioctitos en un ángulo obtuso aproximadamente cercano a los 90º, el verde parece azul y el azul parece púrpura. Por tanto, el método correcto de examen es iluminar el acuario desde el frente. El criterio correcto incluye la clasificación de cualquier espécimen que muestre verde en cualquier ángulo, el verde también puede parecer azul con otra iluminación, pero el azul nunca parecerá verde. El azul acero no es confundido con facilidad con el verde o el azul eléctrico (royal blue). En peces con pocas o ninguna guanáfora en cuerpo y aletas, el reflejo de los iridioctitos encargados de producir el color puede ser fácilmente detectado en la base del ojo.
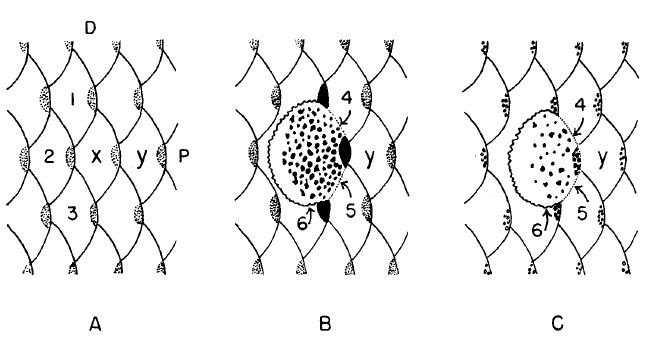
En aras de obviar las dificultades propiciadas por el movimiento de los gránulos de pigmentación para clasificar el color, es preferible hacer un recuento de células de pigmentación. Si las escamas (de la fig 2) son eliminadas del cuerpo, la cobertura de la dermis y epidermis en la porción libre (posterior) permanece intacta.

Estas capas sobre la parte anterior del cuerpo quedan bajo las escamas más próximas. La línea en la cual la separación de la dermis está es muy precisa y corresponde a la línea en la cual las escamas emergen de los estratos de las células. Goodrich and Smith (1937) haciendo un recuento de cromatóforos en el Pez Paraíso designaron la dermis con sus cromatóforos que quedan eliminados sobre las escamas del cuerpo, la zona superficial. La dermis que recubre el cuerpo es divisible en zonas intermedias y profundas. En el Betta la zona intermedia está tan fuertemente pigmentada con la corola o largos tipos interrumpidos de melanóforos (Goodrich 1944) que en la mayoría de fenotipos las células individuales no pueden ser deshechasy la presencia de xanthóforas y erythróforas no puede ser detectada.
La zona profunda también está delimitada, posteriormente por la fuertemente pigmentada capa intermedia, dorsal y ventralmente por pliegues de piel por donde emergen las escamas y anteriormente por la ausencia de los cromatóforos muy por debajo de la próxima zona anterior intermedia. De este modo, antes de cada escama hay una zona profunda de cromatóforos que pueden ser usados como unidades de área.
Todos los recuentos fueron hechos en peces adultos, las áreas medidas no fueron usadas y los cromatóforos de las zonas totalmente profundas fueron contados en orden para eliminar diferencias trazables para heterogeneidades contenidas en ese área.
Para eliminar las diferencias en el número de cromatóforos por unidad de área que dependen de su posición en el pez, la quinta columna de escamas contadas ventralmente en la parte anterior de la aleta dorsal fue adoptado como la región estándar para recuento (escama etiquetada como x en la fig1).
Resultados y discusión:
Un fenotipo de cuerpo claro o grupo de fenotipos conocidos como Camboya han sido trazados hasta un único gen recesivo por Goodrich & Mercer (1934) e independientemente por Umrath (1939) . Afortunadamente, en ambos trabajos el recesivo fue llamado c y el alelo dominante fue denominado
C. El trabajo de Eberhardt (1941) y Wallbrunn (1948) corroboró los hallazgos. Es importante, sin embargo, aclarar la naturaleza de la acción de este gen desde los primeros ensayos, condición que produjo un “Unvollstaendige Albino” o un “pez de cuerpo rosa suave”.
Los machos Camboya son ocasionalmente como el rojo al igual que cualquier pez
C con la única diferencia de la supresión del negro tanto en cuerpo como en aletas. Los Camboyanos no son albinos ya que los ojos están completamente pigmentados con melanina. Los auténticos albinos son conocidos en el Betta pero son ciegos o próximos a serlo, lo cual hace muy difícil tanto su supervivencia como su cría; por ende, ninguna línea de albinos ha sido establecida y la relación entre albinismo y el Locus
C no ha sido demostrada.
En muchas líneas de Camboyanos surge una acumulación considerable de formas negras en los cuerpos de los adultos, como series muy regulares de puntos. Un minucioso examen los revela como proporcionados por los melanóforos en la zona intermedia pero nunca en la zona profunda y rara vez en la zona superficial. Esta aparición negra en los Camboyanos es aparentemente heredada pero no ha sido investigada por su irregular promedio de obtención, a veces desarrollándose sólo después de la poco avanzada edad de 18 meses.
El Rojo se desarrolla considerablemente tarde en la octogenia de los Camboyanos, tanto que en peces oscuros y hembras Camboya no se desarrolla nunca el rojo en su cuerpo, a pesar de que sus aletas sean tan rojas como las de los machos Camboya.
En 1948, un Betta macho rojo brillante fue comprado por el autor. Fenotípicamente el pez parecía ser intermedio entre
cc y
C_. En un test para el alelismo con
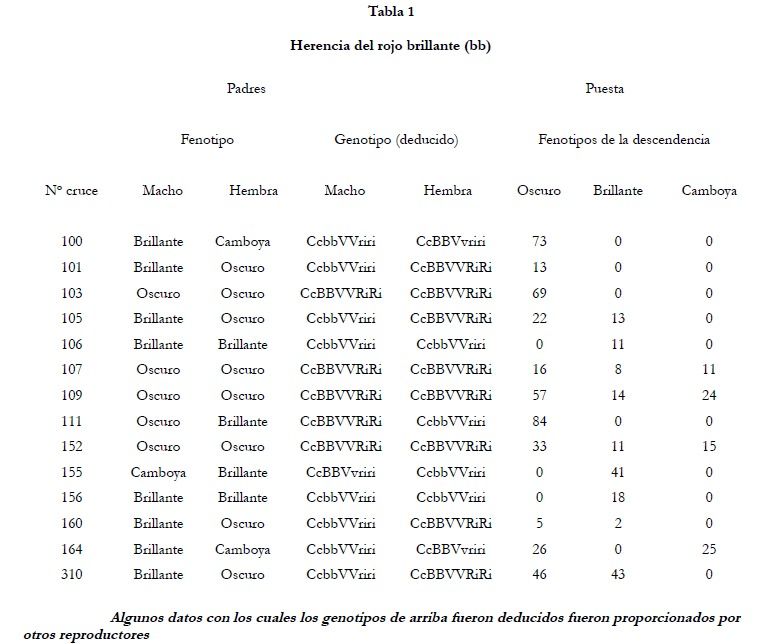
c, el macho fue cruzado con 1 hembra Camboya completamente desprovista de negro en el cuerpo. Su descendencia (puesta # 100) fue totalmente uniforme en el color del cuerpo. 73 peces crecieron hasta la madurez y fueron todos rojo oscuro, más oscuros (contenían más negro) que sus padres (tabla 1).
Hay por tanto, evidencias de un nuevo Locus con 2 alelos, el recesivo del cual los homocigóticos producen el fenotipo rojo brillante, donde esta el
Csuministrado esta presente. El genotipo
cc es epistático para los alelos anteriormente descritos.
El retrocruce (# 105) de una hembra oscura de la puesta # 100 con su padre rojo brillante dio los 2 tonos de rojo en el ratio 2:1 (22 oscuros, 13 brillantes); mientras la F2 (# 107) dio 16 rojo oscuro: 8 rojo brillante: 11 camboyanos. Como las 3 puestas (# 107, # 109 y # 152), en las cuales los 3 colores, rojo oscuro, rojo brillante y Camboya, aparecen de los apareamientos de oscuro x oscuro, son cogidas también, con un resultado de 106:33:49 que es de extrañamente cercano a 9:3:4 (105.8:35.2:47.0). Esto también podría salirse del ratio 2:1:1 pero el cruce # 111, en el que los oscuros se comportan como homocigóticos, no podemos descartar la hipótesis necesaria para el recuento de semejante ratio.
Dentro del grupo rojo brillante, como sucede en el grupo rojo oscuro, hay una gran variedad de tonos, pero las dos clases son muy distintas. El rojo brillante fue usado parea buscar un término mejor para designar la clase, sin embargo algunas de las individualidades recogidas en dichos términos no son especialmente rojas; y el microscopio revela que algunos de los peces oscuros tienen tantos eritróforos y pigmentos como los peces rojo brillante, la diferencia entre las dos clases reside en el número de melanóforos y/o en la acumulación de Melanina presente. El nuevo gen, por tanto, reduce la acumulación de negro y ha sido llamado
b.
B es el alelo que produce la acumulación “normal” de Melanina.
Dentro de la zona profunda de la dermis las diferencias en el número de melanóforos entre
bby
B_ podrían ser muy marcadas como muestran las puestas # 105 y # 107 en la tabla 2. El tamaño de las clases empleadas para hacer el recuento van en incremento, como en series geométricas. Esto se hizo para separar los errores relativos en un recuento ajustado porque los errores absolutos tienen una función en el tamaño del recuento.
En la puesta # 109 la separación de rojos brillantes y rojos oscuros por vía visual fue hecha con facilidad, si bien los recuentos en la zona profunda se acercan a coincidencias. Debe recordarse que hay melanóforos en las otras zonas que también influyen en el color general y éstos también se reducen en número en los peces
bb.
Examinando la tabla 2 se muestra la cercanía entre los alelos
B y
b, el genotipo residual tiene un efecto notable controlando el número de melanóforos en la zona profunda. Existe la posibilidad de que
Bbcause una reducción en comparación con
BB, pero la puesta # 109 muestra que si ese es el caso, el efecto no es demasiado grande para hacer una separación clara del grupo oscuro en 2 clases.
Umrath (1939) describe un fenotipo rojo producido por 1 gen recesivo,
m, “Ein Zuruecktreten der melanophoren” causado por
mm. El gen
m podría ser el mismo que
b pero dado que la descripción de Umrath es fragmentada y sus datos también lo son, no hay seguridad de que los 2 nombres correspondan al mismo locus. Si el color descubierto por Umrath es el mismo que
bb, sería extraño porque más tarde en 1949 los criadores de peces de este campo lo consideraron raro, en vista de ello estas nuevas formas de peces tropicales fueron rápidamente importadas por los criadores.
El promedio de las aletas de peces
C_bbtiene una única línea de melanóforos alrededor del borde exterior. El sobrante de largas áreas restantes en las aletas es densamente cubierto por largos eritróforos pero no por melanóforos. En peces
C_B_ se ve un área típica en la aleta caudal en la cual tienen alrededor de 40 melanóforos por punto bajo el microscopio a alta resolución (430x).
Los azules y verdes cubiertos de iridiscencia por los cuales los peces Betta son famosos son trazables en 2 loci. Uno afecta a la densidad de cobertura de los iridictitos y el otro el espesor de los cristales de guanina y por ende el color particular refractado.
Umrath (1939) llegó a la conclusión correcta de que tanto el verde como el azul oscuro (steel blue) son trazables para los 2 homocigóticos y el azul (royal blue) para el heterocigótico como residuo de un pequeño número de puestas y especímenes. Wallbrunn (1949) corroboró la hipótesis de Umrath y llamó al Locus
G. Eberhardt (1941) también llamó al Locus
V (viridens) pero la guerra mantuvo su papel expansivo en este campo.
VV (=GG) es la designación para el pez azul acero,
vv(=gg) para el verde y
Vv(=Gg) para el azul (royal blue).
En algunos peces los irioctitos son tan escasos que resulta difícil determinar qué color producen. Estos peces son los homocigóticos recesivos, riri (reduzierte Iridocyten (reducción de iridóforos)) (Eberhardt 1941). En algunos fondos genéticos producen manchas iridiscentes en cada escama haciendo hileras de verde, azul y salpicaduras color bronce. Sin embargo, nunca se cumple que el que el homocigótico recesivo tenga extensas áreas cubiertas de iridioctitos. Eberhardt enunció que era posible distinguir
RiRide
ririmediante la acumulación de verde o azul. Sin embargo, no los separó en 2 clases en sus tablas y el presente autor ha tenido varios heterocigóticos tan completamente cubiertos de iridioctitos como cualquier homocigótico. En las notas del autor (Wallbrunn 1948) el dominante fue designado como
S brillante (
Si).
Tests para vinculaciones
Eberhardt (1941) encontró los tres loci
C, Ri y Vpara variar aleatoriamente. Para los tests de vinculación de
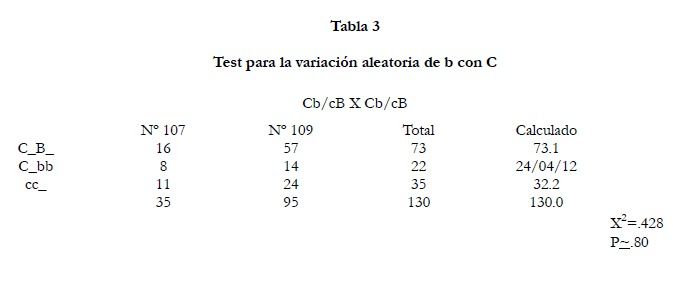
b con
C (tabla 3) sólo las puestas #107 y #109 pudieron ser usadas ya que eran los únicos casos en que estas combinaciones de genes procedentes de los cigotos paternos eran conocidos. Podríamos escribir los genotipos de los padres como fracciones con el significado de que las combinaciones de genes en el numerador o el denominador vengan del mismo gameto. Los cálculos basados en variaciones aleatorias son muy adecuados ya que ninguna vinculación es indicada siempre que el número sea pequeño.
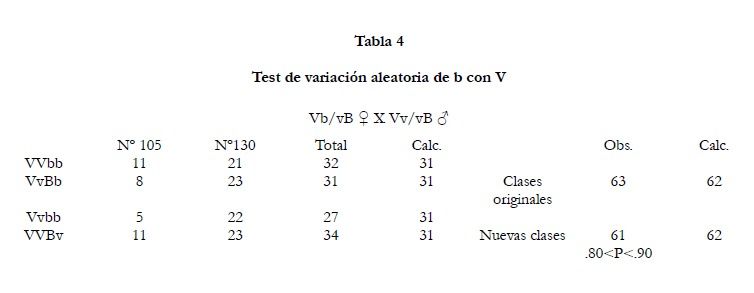
Las puestas # 105 y # 310 pudieron ser usadas para testear las variaciones aleatorias de
b con
V (tabla 4). Los datos combinados están embarazosamente próximos a los valores teóricos calculados para variaciones aleatorias.

La puesta # 310 y el pez
C_ procedente de la puesta # 109 pudieron ser usados para testear la variación aleatoria de
b con ir (tabla 5). Los 2 reproductores son distintos y por tanto son tratados por separado. El pez # 109 obviamente es muy adecuado para la variación aleatoria. Si bien la
Bbririen la puesta # 310 es cuatro veces mayor y la clase
bbRi_ cinco veces más pequeña que las calculadas en las bases de la variación aleatoria. Estos podrían ser más pequeños si los loci están enlazados; por tanto ambos se desvían solo de uno de los valores. Si los datos de las dos puestas son combinados y también las 2 primeras clases y las 2 últimas clases de la tabla combinadas con los 2 primeros podrían ser más largas y las 2 últimas más cortas que lo calculado si
B y
ri están enlazados, los valores observados son 85 y 66 cuando los calculados son 84.4 y 66.6.
Sumario
El amplio rango de colores en el Betta es trazable hasta unos pocos principales y quizás hasta muchos loci menores de los cuales solo unos pocos han sido realmente investigados. La producción y distribución de Melanina está ampliamente controlada por dos loci,
B y
C, ambos dominantes y que deben estar presentes para la producción del común fondo rojo-marrón del cuerpo que es presumiblemente el tipo salvaje. La homocigosis de
b reduce considerablemente el negro en el cuerpo y lo elimina del resto de capas periféricas de células en las aletas. El Camboya es una reducción más drástica del negro en el cuerpo y lo mismo en las aletas como
bb. El gen
c es epistático de
b.
El pez
bbha sido llamado rojo brillante porque siempre parece muy rojizo. Este no es siempre el caso y la acumulación de rojo depende de otros genes los cuales todavía han de ser investigados.
Si la cubierta de irdioctitos está presente da un lustre metálico, el color depende de los alelos en el Locus
V.
Verde = vv, azul real = Vv y azul acero= VV.
Los iridioctitos pueden estar tan dispersos como para hacer también su color indetectable o pueden formar una completa cobertura alrededor del cuerpo y las aletas. Se encuentran todos los grados entre estos dos extremos y generalmente esta cobertura no es uniforme. Un Locus principal (
Ri) controla la densidad dando pocos
ririo no visibles y
Ri_ una considerable acumulación ir es completamente dominante (doble dominante) o próximo a serlo. Lejos de ello, no han sido detectado vínculos en el Betta.

 ¡Leer más!
¡Leer más!